پوشش سلولی ممکن است به صورت غشای سلول و دیواره سلول و در صورت وجود، غشای خارجی به همراه آن ها تعریف شود. دیواره سلولی شامل لایه پپتایدوگلایکان و ساختارهای متصل به آن می شود. اغلب پوشش های سلولی باکتریایی به دو گروه تقسیم می شوند: گرم مثبت و گرم منفی.
این تقسیم بندی بر اساس خصوصیات پوشش در رنگ آمیزی گرم می باشد که نشان دهنده تفاوت های ساختاری مهم بین این دو گروه می باشد. انواع دیگر دیواره سلولی هم در تعداد کمی از گونه های باکتریایی یافت شده اند (نه گرم مثبتند و نه گرم منفی(.
پپتایدوگلایکان، یک ماکرو مولکول بسیار بزرگ پاکتی شکل با اتصالات متقاطع فراوان است که غشای باکتری را احاطه می کند و موجب استحکام و سختی دیواره می شود.
پپتایدوگلایکان شامل اسکلت گلایکان (پلی ساکارید) است که حاوی N-استیل گلوکزامین و زنجیره های جانبی پپتیدی حاوی آمینواسیدهای D و L و در بعضی موارد دی آمینوپایملیک اسید می باشند. زنجیره های جانبی به پل های پپتیدی متصل هستند. این پل ها دارای تنوع ساختمانی در میان گونه های باکتری ها هستند. مورامیک اسید،آمینو اسیدهای نوع D و دی پایملیک اسید توسط پستانداران سنتز نمی شود. پپتایدوگلایکان در همه باکتری ها به جز کلامیدیا و مایکوپلاسما وجود دارد.
پوشش سلولی باکتری های گرم مثبت:
شامل تیکوئیک اسید (پلیمر ریبیتول یا گلیسرول حاوی فسفر) و یا تیکورونیک اسید(پلی ساکاریدهای حاوی گلوکورونیک اسید) می باشد که به شکل کووالان به پپتایدوگلایکان متصلند. عقیده بر این است که این مولکول های دارای بار منفی، در حفظ یون های فلزی نقش دارند.
تیکوئیک اسید، هم چنین می تواند آنزیم های اتولیتیک را به جایگاه هدفشان برای هضم پپتایدوگلایکان هدایت کند (اتولیز) که یکی ار مراحل بیو سنتز دیواره سلولی است. در بعضی موارد نیز پلی ساکاریدهای خنثی وجود دارند. لیپو تیکوئیک اسید، در بسیاری از باکتری ها با غشای سلولی مرتبط است. در موارد دیگر، فیمبریه در خارج سلول شکل می گیرد.
پوشش سلولی باکتری های گرم منفی:
حاوی لیپوپروتئین براون که به صورت کووالان به پپتایدوگلایکان متصل است و هم چنین به غشای خارجی نیز متصل است. مثل دیگر غشاها،غشای خارجی حاوی پروتئینها و پلی ساکاریدها می باشد. اما برخلاف بقیه غشاها، حاوی مولکول های اضافی می باشد (لیپوپلی ساکارید). لیپوساکاریدها موجب ایجاد سد نفوذپذیری به مواد هیدروفوب می شوند.
لیپو پلی ساکارید پامل سه بخش است:آنتی ژن خارجی O،هسته مرکزی و لیپید A در داخل.
هسته مرکزی حاوی چند مولکول قندی است که در طبیعت دیده نمی شود و لیپید A حاوی اسیدهای چرب بتا هیدروکسی است (غیرمعمول در طبیعت). این مولکول دارای فعالیت اندوتوکسیک است. پورین ها در غشای خارجی به ایجاد کانال جهت عبور مواد غذایی کوچک هیدروفیل (مثل قندها) از غشای خارجی کمک می کنند.
باکتری های اسید فست و باکتری های مرتبط (مایکو باکتریا، نوکاردیا و کورینه باکتریا (:
پوشش سلولی این ارگانیسم ها به میزان قابل توجهی پیچیده تر از سایر باکتری هاست. مایکولیک اسید (اسیدهای چرب طویل و شاخه دار) به طور کووالان از طریق یک پلی ساکارید به پپتایدوگلایکان متصل است. دیگر ترکیبات حاوی مایکولیک اسید و لیپیدهای پیچیده دیگر، یک لایه غشایی مومی شکل ضخیم را در خارج از لایه پپتایدوگلایکان تشکیل می دهند.
سنتز ماکرومولکولهای پوشش سلول باکتری:
پپتایدوگلایکان: زیر واحد پیش ساز (مورامیل پنتا پپتید متصل به یوریدین دی فسفاتUDP)در سیتوپلاسم سنتز می شود و به غشای سلول منتقل می شود.
این زیر واحد به شکل آنزیمی از نوکلئوتید به یک ناقل لیپیدی (آندکارپنول/باکتوپرنول) منتقل می شود و به شکل زیر واحد کامل(دی ساکارید پنتا پپتید به همراه پل پپتیدی متصل به آن) ساخته می شود. سپس، زیرواحدهای کامل شده به دیواره سلولی صادر می شوند. پس از رها شدن منومر، آندکاپرنول درون غشا منتشر شده و مصرف می شود.
اسکلت گلایکان دیواره سلولی به شکل آنزیمی شکسته شده (توسط اتولیزین ها) تا امکان ورود زیر واحدهای تازه ساخته شده بوجود بیاید.
اگر این آنزیم ها بیش از حد فعال شوند، دیواره سلولی تجزیه می شود و فشار اسمزی بالای سلول موجب ترکیدن غشای سیتوپلاسمی و مرگ سلول (اتولیز) می شود.
اتصال متقاطع زنجیره جانبی پپتیدی زیر واحد وارد شده به زنجیره موجود به صورت آنزیمی صورت می گیرد (پروتئین های متصل شونده به پنی سیلین). هم چنین زیر واحدهای کامل شده تیکوئیک و یا تیکورونیک اسید هم قبل از انتقال و ورود به دیواره سلولی در غشای سلولی سنتز می شوند.
لیپوپلی ساکارید:
لیپید A در داخل غشای سلولی قرارگرفته و قندهای مرکزی به ترتیب به آن متصلند. زیر واحدهای آنتی ژن O به طور مستقل ساخته می شوند (روی یک حامل لیپیدی). سپس ، آنتی ژن O کاملا ساخته شده قبل از ورود به غشای خارجی به مجموعه لیپیدA-قند مرکزی در غشای سلولی متصل می شود.
اندوسپور: این سلول های باکتریایی گرم مثبت تغییر یافته، دارای پوشش سلولی غیر معمول هستند که شامل یک غشای سلولی و یک غشای خارجی می باشد. لایه پپتایدوگلایکان در این حالت، اتصالات متقاطع کمتری دارد و حاوی شکل دهیدراته مورامیک اسید است. پپتایدوگلایکان اسپور با نام کورتکس مطرح می شود که بین دو غشا قرار دارد. پوششی حاوی مقدار زیادی کراتین با اتصالات متقاطع فراوان در قسمت خارجی سلول وجود دارد. اسپور باکتریایی به دلیل داشتن این پوشش، فوق العاده به عوامل شیمیایی مقاوم است.
به طور طبیعی در تقسیم سلول باکتری، همزمان با تقسیم شدن سلول دیواره ای شکل می گیرد که سلول مادر را به دو سلول دختری هم اندازه تقسیم می کند. در زمان تشکیل اسپور، تقسیم سلول به شکل نا مساوی صورت می گیرد و سلول مادری بزرگتر، سلول دختری را می پوشاند. غشای سلولی سلول دختری ، غشای داخلی اسپور را تشکیل می دهد و غشای سلولی سلول مادری ، غشای خارجی اسپور را تشکیل می دهد
درمقطع ساقه هاي چوبي گياهان دولپه اي ومخروط داران ونيز گياهان ساكن مناطق معتدل،حلقه هاي رشد ديده مي شود اين حلقه ها در مقطع ساقه تك لپه اي ها(خرما ونارگيل)وگياهاني كه در مناطق گرمسير مي رويند(بعلت ادامه رشد در طول سال) ديده نمي شوند .
در اين گياهان سال به سال به علت زياد شدن آوند ها قطر ساقه زيادتر مي شود.
به طوري كه با برش ساقه گياه حلقه هاي متحدالمركزي ديده مي شود كه به آنها حلقه هاي رشد سالانه مي گويند.(لايه هاي ضخيم چوب پسين يا چوب اغلب حلقه اي شكل هستند)با برش حلقه هاي عرضي و مشاهده ي رشد سالانه گياه مي توان به شرايط گذشته زندگي گياه پي برد (حلقه های سالانه نه تنها سن درخت را نشان می دهند بلکه آنها می توانند شرایط محیطی و اقلیمی را که گیاه در طول زندگی خود گذرانیده است به ما بازگو نمایند . )
حلقه هاي سالانه در واقع آوند هاي چوبي از كار افتاده ي مربوط به سال هاي قبل اند اگر با ذره بين به مقطع صيقل شده ي يك ساقه ي چند ساله نگاه كنيم دهانه ي آوند ها را به صورت لانه ي زنبور مشاهده خواهيم كرد .هر حلقه كه نتيجه ي فعاليت يك سال كامبيوم است، از دوقسمت تيره وروشن تشكيل شده است در قسمت هاي روشن ، آوند ها بازتر بوده وآب واملاح بيش تري را منتقل مي كرده انديعني دراين قسمت هاياخته ها بزرگتر وديواره آن ها نازكتر است(در بهار) در قسمت هاي تيره، دهانه ي آوند ها كوچك تر است وآوند ها به صورت فشرده به هم قرار دارند(درتابستان و پاييز)
نكته: هر سال تنها یک حلقه رشد می کند
سن درخت را مي توان با شمارش حلقه ها در سطح مقطع تنه درخت(در نزديكي خاك)تعيين كرد.وقوع حلقه هاي رشد كاذب موجب مي شود كه سن درخت زيادتر تخمين زده شود.ازجمله عوامل مسئول در توليد حلقه هاي كاذب خشكي وبي برگ شدن گياه بوسيله حشرات است.
ضخامت حلقه ها معمولا مساوي نيست وبستگي به عوامل محيطي مانند نور،دما،باران ،رطوبت،خاك فطول فصل رويش ورقابت دارد .بديهي است هرچه فعاليت گياه در طول يك سال بيش ترباشد،قطر حلقه هاي مربوط به آن سال هم بيش تر خواهد بود.
برای تعیین سن درخت ، نیاز به بریدن آن نیست . گیاه شناسان و جنگل بانان از دستگاههای سوراخ کننده ای برای این منظور استفاده می کنند . در این شیوه یک قطعه میله ای شکل از چوب درخت را عمود بر تنه آن برمی دارند . حلقه های سالانه سپس در این قطعه چوب شمارش می شود . سوراخ کوچک باقیمانده روی تنه درخت را پس از آن با مواد ضدعفونی کننده پرمی کنند تا از ورود بیماری به آن جلوگیری شده و به درخت آسیبی وارد نشود .
حلقه هاي ساقه گياه دوساله چوبي(ازداخل به خارج)
پارانشيم مغزي - چوب نخستين- چوب سال اول- چوب سال دوم- كامبيوم آوندي- آبكش سال دوم- آبكش سال اول- آبكش نخستين- پوست - كامبيوم چوب پنبه ساز- چوب پنبه
رشد در گیاهان بر خلاف جانوران ، بطور یکنواخت اتفاق نمیافتد. زیرا در گیاهان رشد به مناطق خاصی محدود میشود. در گیاهان دو نوع رشد ، تحت عنوان رشد اولیه و رشد ثانویه تشخیص داده شده است. رشد اولیه در انتهای اندامهای هوایی یا ریشهها و در زواید جانبی مانند برگها و جوانهها رخ میدهد. از آنجا که رشد اولیه در نوک ریشهها و اندامهای هوایی اتفاق میافتد، گیاهان میتوانند دارای رشد نامحدود باشند.
در برخی از گیاهان مانند ذرت و آفتابگردان ، گلدهی پس از تولید تعداد مشخصی برگ صورت میگیرد و پس از گلدهی رشد رویشی متوقف میشود. در بیشتر گیاهان بجز تک لپهایهای درختی ، رشد اولیه ضرورتا معادل رشد طولی است. ساقهها و ریشه در بسیاری از گیاهان دارای رشد قطری نیز میباشند که به این افزایش قطر ، رشد ثانویه اطلاق میشود. رشد ثانویه در ناحیهای رخ میدهد که رشد طولی متوقف شده باشد.
بافت مریستمی
رشد طولی دارای مناطقی است که سلولها در حال رشد سریع هستند. این بافتهای دائما زایا ، مریستم نامیده میشوند. سلولهای بافتهای مریستمی دارای دیواره نازک ، هسته بزرگ و در اکثر موارد واکوئلها کوچک هستند. مریستمهای انتهایی در انتهایی ریشهها و اندامهای هوایی وجود دارند و منجر به افزایش رشد اولیه و یا رشد طولی میشوند. همه سلولهای مریستمی دارای قدرت تکثیر هستند و پس از تکثیر میتوانند رشد کرده، تمایز یافته، با تغییر شکل ، بافتهای مختلف را بوجود آورند.
ریشههای اولیه را میتوان به 4 منطقه تقسیم بندی نمود
ریشهها معمولا اولین اندامی هستند که طی جوانه زنی بذر پدیدار میشوند. معمولا ریشه به 4 منطقه تقسیم میشود که عبارتند از: کلاهک ریشه ، مریستم انتهایی ، منطقه طویل شدن و منطقه تارهای کشنده یا منطقه بلوغ. البته بجز کلاهک ریشه ، مرز این مناطق تا حد زیادی با یکدیگر همپوشانی دارد. سلولهای مریستم انتهایی همزمان با اینکه تقسیم میشوند، توسعه مییابند و بنابراین در منطقه طویل شدن قرار میگیرند. بسیاری از سلولهای واقع در منطقه طویل شدن در حال تمایز یافتن هستند و سلولهای خاصی مانند تارهای کشنده در منطقه بلوغ طویل میشوند.
کلاهک ریشه
سلولهای کلاهک ریشه از یک مریستم خاص کلاهک ریشه بوجود میآیند و سلولهای قدیمیتر به تدریج به طرف نوک ریشه منتقل میشوند. همینطور که سلولهای کلاهک بالغ میشوند، عمل تمایز نیز در آنها صورت میگیرد. و هنگامی که کاملا تمایز یافتند نسبت به نیروی جاذبه زمین حساس میشوند و کلاهک ریشه غالبا باعث حفاظت مریستم انتهایی از صدمات مکانیکی میشود.
مریستم انتهایی
تقسیمات سلولی انجام شده در مریستم انتهایی ، منشا همه سلولهای ریشه است. هیچگونه زایده جانبی نزدیک منطقه انتهایی ریشه تشکیل نمیشود. سلولهای مریستم انتهایی کوچک و تقریبا مکعبی شکل هستند. این سلولها تا مدتی تقسیم میشوند.
منطقه طویل شدن
این منطقه ، محلی است که رشد ریشه سریع است و سلولها ضمن اینکه توسعه مییابند دائما تقسیم میشوند.
منطقه بلوغ
پس از اینکه تقسیم و طویل شدن سلولها متوقف شد، سلولها بالغ شده، و ستونی از انواع مختلف سلول را تشکیل میدهند. گسترش ریشههای فرعی در این منطقه ، شاخص خوبی برای نشان دادن پایان منطقه طویل شدن است. در منطقه بلوغ ، استوانه مرکزی بافت آوندی ، پس از تشکیل توسط آندودرم احاطه میشود و خود این لایه توسط لایهای از سلولهای پوست محصور میگردد.
رشد طولی ساقه
رشد ساقه نیز مانند رشد ریشه ، حاصل تولید مثل سلولهای مریستم انتهایی است. سلولهایی که در ورای این مریستم تولید میشوند، ابتدا طویل شده و نوک ریشه در حال رشد را به سمت بالا میرانند و آنگاه به شکل بافتهای متحدالمرکز ساقه کامل ، تمایز مییابند. مریستم راس ساقه بوسیله کلاهکی از سلولها پوشانده نشده است، اما از جهات دیگر ، بافتهای ریشه و ساقه بهم شباهت دارند. ساقه از نمو جوانه انتهایی و شاخهها از نمو جوانههای جانبی یا نابجا بوجود میآیِند. منطقه رشد طولی که زیر انتهایی است و با عدم تساوی طول میانگرهها قابل تشخیص است. رشد ساقه با رشد میانگرهها تکمیل میگردد.
رشد برگ
برگها از تولید مثل سلولهایی که در پهلوهای مریستم انتهایی ساقه جای دارند، حاصل میآیند. سلولهای این قسمت ، سریعتر از سلولهای اطراف خود تقسیم میشوند و زایده های کوچک انگشت مانندی به نام جوانه انتهایی برگ را پدید میآورند. این ساختارهای جنینی تدریجا رشد میکنند و به برگهای کامل تبدیل میشوند. نوک ساقهها معمولا دارای تعدادی جوانه اولیه برگ است که فاصلههای میان گرهی بسیار کوتاهی آنها را از همدیگر جدا میسازند.
بیشتر رشد طولی ساقه به علت طویل شدن سلولهایی صورت میگیرد که همین فاصلههای میان گرهی را تشکیل میدهند. اما جوانههای اولیه برگی که فاصله کوتاه میان گرهی را در راس ساقه از هم جدا میکنند، قبل از آنکه طویل شوند در اطراف مریستم انتهایی تجمع مییابند و مجموعهای به نام جوانه انتهایی را میسازند. با آغاز رشد جوانه اولیه برگ ، سلولهای قاعده آن دمبرگ و بقیه سلولها پهنک و در برخی گیاهان گوشوارک را بوجود میآورند.
رشد شاخه ها
پیش از کامل شدن تمایز برگ ، جوانه نوع دومی معمولا در زاویه میان ساقه و دمبرگ رشد میکند. این جوانههای مولد شاخه نظیر همان ساختمان جوانههای انتهایی را دارند و میتوانند مبدل به شاخه شوند. جوانههای مولد شاخه اغلب به سرعت رشد نمیکنند. پارهای از آنها به مدت چندین سال در حال زندگی نهفته میمانند. در فصل زمستان ، جوانههای مولد شاخه غیر فعال ، درست در بالای مناطق مدور و کوچکی که محل اتصال برگها در فصل رشد گذشته بودهاند، مشاهده میشوند. به این نقاط ، اثر برگ گفته میشود.
اثر هورمون اکسین بر رشد طولی گیاهان هورمون اکسین عامل مهم گسترش سلول است. رشد پهنک برگ دو لپهایها تحت تاثیر اکسین ، کاهش دارد. بر عکس پهنک برگ تک لپهایها ، دمبرگ و غلاف برگ دو لپهایها تحت تاثیر اکسین رشد میکنند. رشد طولی ساقه تحت تاثیر اکسین زیاد ، افزایش مییابد. رشد طولی ریشهها بوسیله اکسین زیاد ، کاهش مییابد اما ریشهزایی افزایش مییابد.
اثر هورمون جیبرلین بر رشد طولی گیاهان افزایش میان گرههای ساقه توسط هورمون جیبرلین آندروژن ، باعث ایجاد قد طبیعی در گیاهان میشود. در ارقام کوتوله گیاهان ، اعمال جیبرلین اگزوژن باعث تحریک رشد و رسیدن ارتفاع آن به حالت طبیعی میشود. مطالعات درباره ژنهای متعددی که جهش در آنها عامل کوتولگی است و در بیوسنتز جیبرلین دخالت دارند، انجام شده است.
اثر هورمون اسید آبسیزیک بر رشد طولی گیاهان
این هورمون باعث کاهش رشد اندام هوایی میشود و سطح برگها نیز کاهش مییابد که به تحمل گیاه در شرایط تنش خشکی کمک میکند.
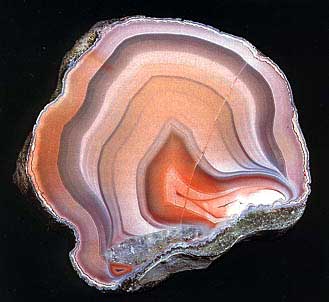
یاقوت یکی از کانیهای سنگهای آذرین، است که بعد از الماس سختترین کانیها محسوب میشود.
یاقوت در رنگهای سرخ و آبی و زرد در دسترس میباشد. در خرافات نشانه قدرت است.
یاقوت سرخ نامش از قرمز گرفته شده ، کروندوم جواهری با کیفیت ، یکی از بهترین سنگهای جواهر است که در جواهر سازی بکار میرود. یاقوت ها به رنگ های مختلف قرمز می توانند باشند، از صورتی تا جگری (زرشکی) یا قرمز قهوه ای پررنگ که بستگی به مقدار آهن و کرومیوم سنگ دارد. تکرار بلورهای مرکب کانی متساعد برای انکسار
می سازند. در استحکام و عدم شکنندگی یاقوت سرخ اولین و از نظر سختی دومین بعد از الماس است. بلورهای منشور 3 وجهی با انتهای مخروطی یا مسطح هستند. هم زمان با رشد بلورها و بسته به شرایط زمین شناختی و کانی های موجود لایه های جدیدی تشکیل می شود و تنوع رنگ ها به نام منطقه بندی رخ می دهد.
كوارتز (sio2)در حالت عادي بي رنگ است .كه به آن در كوهي گفته مي شود. در صورت وجود نا خالصي يه رنگ هاي شيري،قرمز،بنفش. . . . ديده مي شود.
عقيق كوارتز قرمز رنگ است و آمتيست كوارتز بنفش رنگ است
از كوارتز در جواهر سازي ، شيشه سازي ، تهيه كاغذ سمباده ،...... استفاده مي شود
آمتيست نوعي كوارتز است كه بخاطر ناخالصي منگنز چنين رنگ زيبايي پيدا مي كند.
این کانی(الیوین) سبز زیتونی با جلای شیشه ای و فاقد رخ است.جزء کانی حرارت بالاست در مجاورت آبهای گرم به سرپانتین و سپس به تالک تیدیل می شود
28-نورون ها چه اعمالي انجام ميدهند؟(0.75 –نکات علمی
نوشته شده در تاريخ چهار شنبه 16 اسفند 1391برچسب:,
توسط ناصر مستقیمی
|
هواي باقيمانده، به مقدار هوايي گفته مي شود که در عميق ترين بازدم هم از ششها خارج نمي شود و در نتيجه، جزء ظرفيت حياتي نيست.
هواي مرده، بخشي از هواي دمي است که درون مجاري تنفسي را پر مي کند و به ششها وارد نمي شود.
به اين ترتيب، هواي باقيمانده در شش است و هواي مرده در مجاري تنفسي.
هواي باقيمانده در دم و بازدم شرکت نمي کند ولي هواي مرده در دم و بازدم شرکت مي کند.
هواي باقيمانده در تبادلات گازهاي تنفسي با خون شرکت مي کند ولي هواي مرده در تبادلات شرکت نمي کند.
مقدار هواي مرده در حالات مختلف تنفسي يک فرد ثابت و به اندازه حجم مجاري تنفسي فرد است که حدودا يک سوم هواي جاري است؛ يعني يک سوم 500 سي سي، نه يک سوم عمق دم.
گربه همانند انسان دارای دید دوچشمی بوده و اجسام را سه بعدی می بیند با این وجود , چشم گربه که برای ثبت حرکات طراحی شده است تفاوت های زیادی با چشم انسان ها دارد, مردمک چشم گربه برخلاف مردمک چشم ما , گرد نیست بلکه بیضوی شکل است . مردمک های بیضوی شکل نسبت به انواع گرد , خیلی سریعتر باز و بسته شده و موجب می شود که اندازه بزرگتری داشته باشد , به این دلیل یک گربه می تواند مردمک خود را به اندازه قرنیه اش ( تقریباً تا 90 % سطح چشم خود) باز کند پس موجب می شود که نور بیشتری وارد چشم شود . در چشم گربه بر خلاف چشم ما تعداد سلول های استوانه ای نسبت به سلول های مخروطی بسیار بیشتر است , بنابراین چشم گربه برای دیدن اجسام در تاریکی سازگاری بیشتری نسبت به چشم انسان ها دارد . این جانوران رنگ های ارغوانی , آبی , سبز و زرد را می بینند اما نسبت به رنگ قرمز , نارنجی و قهوه ای کوررنگ هستند گربه ها این رنگ ها را بصورت خاکستری می بینند .
نکته جالب درخشش چشمگربه ها در تاریکی است . در پشت شبکیه چشم گربه ها لایه ای از سلول های منعکسکننده نور وجود دارد که tapetum lucidum نامیده می شود این لایه ی آینه مانند , نور تابیده شده به شبکیه را منعکس می کند و موجب می شود که سلول های استوانه ای دوبار در معرض نور قرار گیرند , بنابراین در تاریکی بسیار بهتر از انسان ها می تواندببینند . وجود این لایه به همراه داشتن مردمک بیضی شکل , موجب شده است که گربه هافقط به یک ششم نوری که ما برای دیدن نیاز داریم به وضوح اجسام را ببینند , بطوریکهدر یک شب نسبتاً ابری با تعدادی ستاره , نور لازم برای دیدن گربه فراهم می شود.
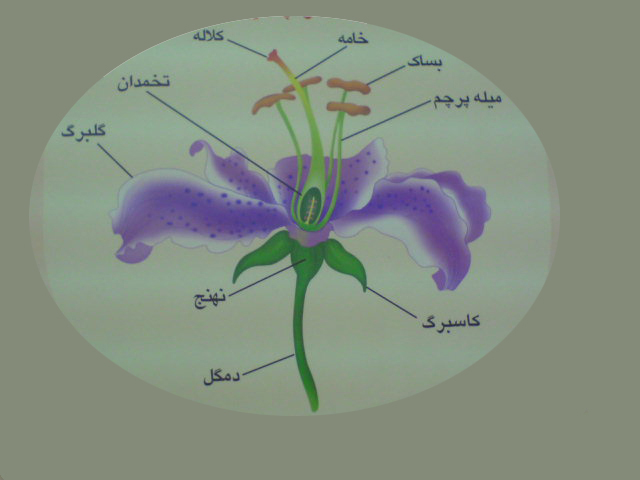
یک گل کامل از چهار حلقه تشکیل شده است که عبارتند از:
کاسبرگ، گلبرگ، پرچم و مادگی .
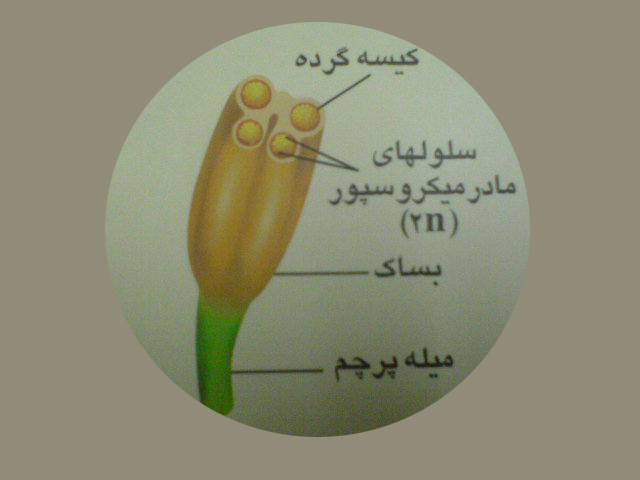
پرچم اندام تولید مثل نر است وشامل دو بخش میله وبساک می باشد .مادگی اندام تولید مثل ماده است. واحد سازنده مادگی ،برچه نام دارد. هر برچه شامل کلاله ، خامه وتخمدان است . در هر بساک رسیده چهار کیسه گرده تشکیل می شود .
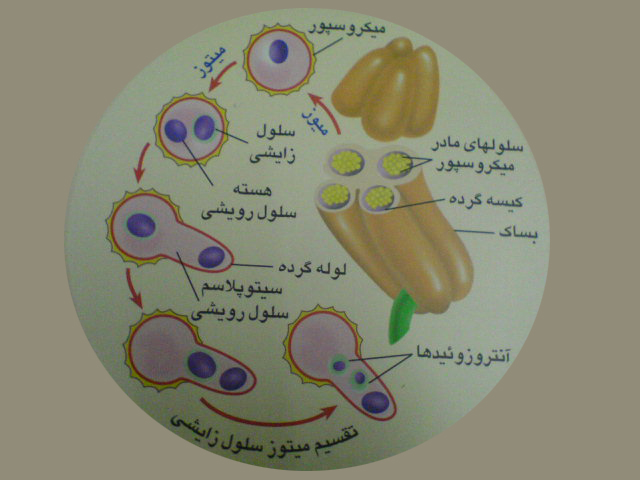
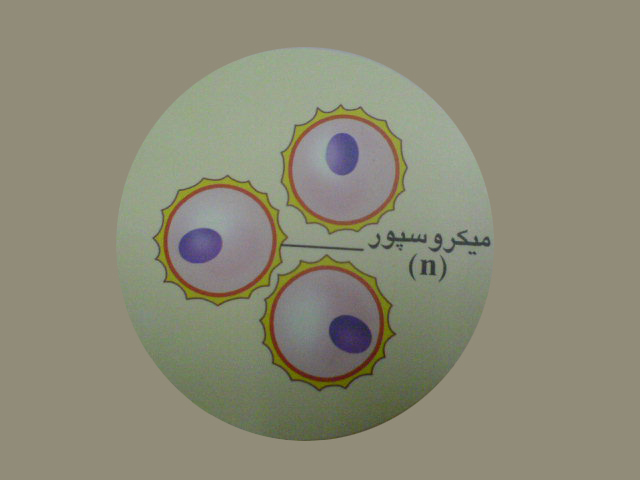
درون کیسه های گرده سلول های مادر میکروسپور دیپلوئیدباتقسیم میوزچهار میکروسپور هاپلوئید را به وجود می آورد . هسته میکروسپور باتقسیم میتوز دو هسته راایجاد کرده که تبدیل به دوسلول رویشی وزایشی می شوند . دانه گرده رسیده دارای دوپوسته خارجی وداخلی نیز می باشد که سلول های رویشی وزایشی رادربرمی گیرند. دانه گرده همان گامتوفیت نر است.
تخمک ها درون تخمدان تشکیل می شوند .تخمک نهاندانگان شامل دوپوسته، سفت وپارانشیم خورش است .
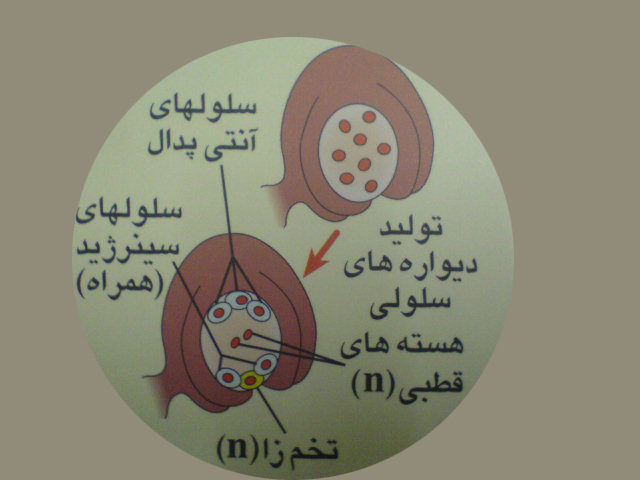
یکی از سلولهای پارانشیم خورش به نام سلول مادر مگاسپور که دیپلوئید است باتقسیم میوز چهار مگاسپور هاپلوئید را به وجود می آورد. سه تااز این سلول ها ازبین رفته، مگاسپور باقی مانده سه میتوز متوالی انجام داده به این ترتیب هشت هسته هاپلوئیدایجاد می شود. هر هسته به یک سلول تکامل می یابد. به این ترتیب کیسه رویانی که گامتوفیت ماده است درون تخمک تشکیل می شود .
در کیسه رویانی هفت سلول مشاهده می شود . سه سلول در قطب مجاور سفت قرار می گیرندکه یکی از آنها تخمزا ودوتای کناری سلول های سینرژید نام دارند.دروسط کیسه رویانی یک سلول درشت بادو هسته هاپلوئید ی به نام سلول دوهسته ای قراردارد. سه سلول دیگر که درقطب مقابل سفت قراردارند سلولهای آنتی پدال نامیده می شوند.
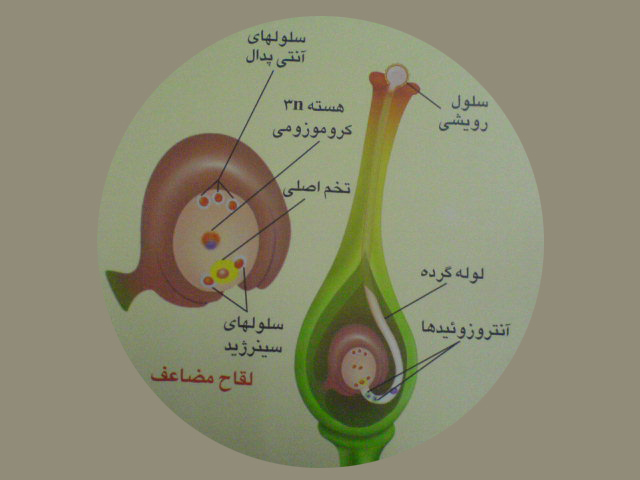
پس از گرده افشانی براثرفعالیت سلول رویشی لوله گرده تشکیل می شود. لوله گرده از سفت عبور کرده وارد تخمک می شود.
لوله گرده از سفت عبور کرده وارد تخمک می شود. سلول زایشی با تقسیم میتوز دو آنتروزوئید را به وجود می آورد.
آنتروزوئید اول باتخمزا لقاح می یابدوسلول تخم دیپلوئیدرابه وجود می آورد.آنتروزوئید دیگر باسلول دوهسته ای لقاح می یابدوسلول تریپلوئیدرابه وجودمی آورد. این نوع لقاح را لقاح مضاعف می گویند. لقاح مضاعف فقط در نهاندانگان انجام می شود .
ازرویش تخم دیپلوئید ،رویان وازرشدسلول تریپلوئید، آلبومن تشکیل می شود .
پس از لقاح ، تخمک تبدیل به دانه می شود . دانه نهاندانگان شامل پوسته ، رویان واندوخته غذایی است .
دانه در شرایط مساعد می روید واز رشد رویان گیاه اسپوروفیت تشکیل می شود .
اختلاف منظر دید ستارگان که ناشی از تغییر موقعیت زمین در فاصله زمانی 6 ماه است. روش اندازهگیری آن به این شکل است که از ستاره موردنظر عکسبرداری می شود، سپس از همان ستاره در فاصله 6 ماه دیگر که زمین به اندازه قطر مدارش حول خورشید جابجا شده است مجدداً عکسبرداری میشود. مکان ستاره در دو عکس مقداری جابجایی نشان می دهد که این جابجایی ظاهری همان اختلاف منظر سالیانه آن ستاره می باشد.مقدار جابجایی برابر با زاویه جدایی بین زمین وخورشید از دید آن ستاره است.این کمیت با نام اختلاف منظر خورشید مرکزی(heliocentric parallax)هم شناخته می شود. از این کمیت به کمک روابط مثلثاتی در تعیین فاصله ستارگان استفاده میشود.
علاوه بر این حرکت ظاهری ،ستارگان دارای یک حرکت دیگر هم هستند که ناشی از تغییر مکان آنها نسبت به خورشید می باشد(حرکت نسبی).
تاسال ۲۰۰۲ فاصله حدود ۴۰۰ ستاره با دقت یک درصد با این روش تعیین شده وفاصله حدود ۷۰۰ ستاره هم با دقت ۵ درصد تعیین شده است.فاصله ستاره هایی که کمتر از ۲۰۰ پارسک باشند با دقت خوبی با این روش قابل محاسبه است.گفتنی است حداقل اختلاف منظر قابل محاسبه در حدود یک هزارم ثانیه قوسی است.
اختلاف منظر بهمراه چندین پارامتر دیگر قابل محاسبه در فهرستهای ستاره ای مانند فهرست تیکو ثبت شده است.
چرا ماه به روی زمین سقوط نمی کند؟
زمین با نیروی گرانش خود ماه را به سوی خود می کشد. اگر انسان ماه را که در حقیقت بی وقفه به دور سیاره ما می چرخد از گردش باز می داشت ماه فقط برای مدت کوتاهی ثابت می ایستاد آنگاه با سرعتی فزاینده به سمت زمین می شتافت و در نهایت با آن برخورد می نمود. البته این عمل میسر نیست. ماه از همان زمانهای اولیه با سرعتی برابر 3659 کیلومتر در ساعت به دور زمین در حال گردش بوده است. در اثر این حرکت گردشی یک نیروی گریز از مرکز به سمت خارج ایجاد می شود که درست به اندازه نیروی گرانش زمین که به سمت داخل کشش دارد است. این دو نیروی مخالف اثر یکدیگر را به طور متقابل خنثی می کنند به نحوی که ماه همواره بر مدار خود باقی می ماند.
منظور از دریاهای ماه چیست؟
لکه هایی که با چشم عادی و غیر مسلح روی سطح ماه مشاهده می شود قبلا" به شکل اقیانوس به نظر می رسید و برای همین دریا نام داده شد. امروزه می دانیم که نیروی جاذبه ماه آنقدر کم است که قادر به نگاه داشتن آب و هوا نیست. بنابراین ماه هیچگاه دارای اقیانوس نبوده است. دریاهای ماه سطوح عظیم و کاملا" خشک از سنگ گدازه ها هستند. این گدازه ها میلیاردها سال پیش در بخشهایی از سطح سفید ماه که مملو از دهانه های آتشفشانی بوده سرازیر شده و روی قسمتهایی از آنها را پوشانیده است. سپس این حجم عظیم گدازه ها سرد شده اند. در حال حاضر چون مانند سابق اجرام آسمانی زیادی در فضا وجود ندارد دریاهای ماه به ندرت مورد اصابت گلوله های آتشین و بزرگ فضایی واقع می شوند به طوریکه این مناطق تاریک دارای دهانه های آتشفشانی بسیار کمتری نسبت به چشم اندازهای سفید قدیمی هستند.
کوتوله های سفید و ستارگان نوترونی چه هستند؟
ستارگان پیر و قدیمی مانن خورشید ما به تدریج لایه های بیرونی خود را دفع می کنند. ستارگان سنگین در پایان عمر خود منفجر می شوند. البته اکثر اوقات یک هسته تمام سوخته از آنها باقی می ماند. اگر این هسته کمتر از 4/1 جرم خورشید وزن داشته باشد به یک کوتوله سفید تبدیل می شود. یعنی کره ای که نزدیک به اندازه کره زمین است. البته در این کره کوچک تمام توده جرم خورشید متراکم می شود به نحوی که یک قاشق چای خوری از ماده کوتوله سفید چندین تن وزن خواهد داشت.
شگفت تر از این سرنوشت باقیمانده ستارگانی است که بین 4/1 تا 3 برابر جرم خورشید وزن دارند. آنها به صورت کره هایی که حدود 20 کیلومتر قطر دارند فرو می پاشند. یک سانتیمتر مکعب از مواد آنها شاید چیزی حدود 100 میلیون تن وزن خواهد داشت. این باقیمانده های ستاره ای را با نام ستاره های نوترونی مشخص می کنند. زیرا آنها تقریبا" به طور کامل از ذراتی ساخته شده اند که نوترون نام دارند. انسان می تواند ستاره های نوترونی زیادی را در آسمان نظاره کند. این کره های کوچک ولی پر جرم بسیار سریع حول محور خود می چرخند. روی این کره های کوچک نقاطی وجود دارد که پرتو افشانی بسیار زیادی دارند. هرگاه این نقاط به طرف زمین قرار می گیرند نوری شدید و لحظه ای دریافت می شود. در واقع انسان در آسمان یک منبع نور و تشعشع تپشی یا لحظه ای مشاهده می کند. به این جهت ستارگان نوترونی را تپ اختر نیز می نامند.
سیاهچاله چیست؟
کوتوله های سفید و ستاره های نوترونی را همواره می توان دید. اما مشاهده باقیمانده های ستاره ای که وزنی بیش از 3 برابر جرم خورشید دارند امکانپذیر نیست. این ستاره ها به صورت ساختارهای چنان کوچک و متراکمی فرو می پاشند که بر سطح آنها نیروی گرانش غیر قابل تصوری حکمفرما می شود. این نیروی گرانش در نهایت چنان زیاد می شود که همه چیز حتی نور را جذب می کند و در خود به دام می اندازد. چنین باقیمانده های ستاره ای را به همین دلیل نمی توان دید و لذا آنها را سیاهچاله می نامند. این ساختارهای شگفت انگیز همچون گردابی عظیم تمام چیزهایی را که نزدیک آنها می شوند ببلعند اما نمی گذارند هیچ چیز حتی نور از دست آنها رها شود و بگریزد.
چون سیاهچاله ها هیچ پرتویی از خود نمی تابانند نمی توان آنها را به طور مستقیم مشاهده کرد. با این وجود اخترشناسان بر این باورند که قراین و شواهدی برای اثبات وجود این اجرام آسمانی یافته اند. برای مثال ستارگانی را می توان در آسمان یافت که دور مرکزی ناپیدا در گردشند. که این مرکز در واقع فقط می تواند یک سیاهچاله باشد. سیاهچاله گاهگاه موادی را از این ستاره می رباید که در این فرایند این مواد شدیدا" داغ می شوند و قبل از آن که توسط سیاهچاله بلعیده شوند پرتوهای رونتگن (اشعه ایکس) از خود می تابانند. این پرتوها را آوای مرگ ماده نیز می نامند.
چرا ما از روی زمین فقط نیمی از ماه را می بینیم؟
ماه تقریبا" 27 روز طول می کشد تا یک بار به دور زمین بگردد. در همین زمان ماه دقیقا" یک دور کامل نیز به دور خود می چرخد. به این جهت ماه همواره یک روی خود را به ما نشان می دهد. انسان از روی زمین فقط یک روی کره ماه را می تواند ببیند. پشت ماه را فقط فضانوردی که با سفینه خود به دور ماه می چرخد قادر است مشاهده کند. به هر جهت انسان می تواند به دلیل نامنظمی های گوناگون در مدار ماه و نیز از طریق تغییر مکان مشاهده ماه در روی زمین در طول زمان 59% سطح ماه را مشاهده کند. بقیه 41% سطح ماه قبل از آغاز دوران سفر به فضا برای انسان کاملا" ناشناخته بود.
منظور از قدر مطلق ستارگان چیست؟ برای مقایسه درخشندگی واقعی ستارگان باید قدر مطلق آنها را تعیین کرد. قدر مطلق عبارت است از قدر ظاهری ستاره هنگامی که فاصله آن با ما 10 پارسک (6/32 سال نوری) باشد.
برای مثال اگر خورشید 10 پارسک دورتر از ما بود همانند ستاره ای کم نور با قدر ظاهری 8/4 دیده می شد. قدر مطلق شعرای یمانی 5/1+ است . زیرا آن هم نزدیکتر از فاصله استاندارد قرار دارد. دنب نورانیترین ستاره صورت فلکی دجاجه از قدر اول است ولی قدر مطلق آن نشان می دهد که یکی از درخشانترین ستارگان راه شیری است. زیرا اگر در فاصله استاندارد قرار می گرفت همانند ستاره ای با قدر 7- دیده می شد. در این صورت به غیر از ماه و خورشید نورانیترین جرم آسمانی بود و حتی نور آن می توانست سایه ایجاد کند.
تفاوت بین دنباله دارها و شهابها چیست؟
گاهی مردم دنباله دارها را با شهابها اشتباه می گیرند در حالی که این دو کاملا" مجزا از یکدیگرند. شهابها در واقع ذرات بسیار ریزی هستند که در فضای میان سیارات پراکنده اند. هنگامی که این ذرات به جو زمین وارد می شوند بر اثر اصطکاک با جو می سوزند و به صورت ردهای نورانی دیده می شوند. ولی دنباله دارها توده هایی از یخ و غبار و ذرات ریز هستند که در مدارهای طولانی به دور خورشید می گردند. اندازه هسته اصلی دنباله دارها بزرگتر از چند کیلومتر نیست. اما هنگامی که به نزدیکی خورشید می رسند به دلیل تابش شدید خورشید گاز و غبار آنها جدا می شود و دنباله ای خلاف جهت تابش خورشید تشکیل می شود که ممکن است طول آن به میلیونها کیلومتر برسد.
صورت فلکی چیست؟
صورت فلکی گروهی از ستارگان در آسمان هستند که مجموعه های قابل تشخیص را تشکیل می دهند. ستاره شناسان قدیمی توانستند بعضی از این مجموعه ها را پیدا کنند و آنها را به نام موجودات افسانه ای و خدایان و الهه ها نامگذاری کنند. امروزه 88 صورت فلکی شناخته شده است و کل آسمان طوری تقسیم شده است که هر ستاره به یک صورت فلکی متعلق باشد. اما ستارگان موجود در هر صورت فلکی چندان ارتباطی با هم ندارند. فقط طوری قرار گرفته اند که وقتی از زمین به آنها نگاه می کنید در یک مجموعه قرار دارند. بدیهی است چنانچه از یک نقطه دیگر فضا به آنها نگاه کنیم مجموعه ها به صورت دیگری به نظر می آیند. در بسیاری از موارد فاصله ستارگانی که یک صورت فلکی را تشکیل می دهند از یکدیگر بیش از فاصله ای است که با ما دارند.
سحابی چیست؟
در بسیاری از مناطق فضای میان ستاره ای ابرهای بزرگی از گاز و غبار وجود دارند که آنها را سحابی (به معنای ابر) می نامند. سحابیها را به سه گروه رده بندی می کنند: سحابیهای نشری- بازتابی و تاریک.
در سحابیهای نشری یک یا چندین ستاره بسیار سوزان (از رده های طیفی Oیا B) جا دارند. این ستاره های بسیار داغ موجب تحریک گازها و درخشش سحابی می شوند. نمونه جالب توجهی از این گونه سحابی بزرگ جبار است. این سحابی با چشم غیر مسلح به صورت توده مه آلود کم نوری دیده می شود. اگر ستاره ها مقداری سردتر باشند یا اینکه چگالی گازها در سحابی زیاد باشد گازها فقط نور ستاره ها را بازتاب می دهند. در این صورت سحابی را بازتابی می نامند. سحابی که ستاره های خوشه پروین را در بر گرفته از نوع بازتابی است.
در برخی موارد هم هیچ گونه ستاره ای در درون یا نزدیکی سحابی قرار ندارد. به همین جهت سحابی را تاریک می نامند. مشاهده سحابیهای تاریک فقط در صورتی ممکن است که در مقابل سحابیهای نشری یا بازتابی قرار گیرند. سحابیهای تاریک نور ستاره های پشت خود را جذب می کنند. اخترشناسان عقیده دارند که ستاره ها در درون این سحابیها متولد می شوند. سحابی سر اسبی نمونه جالب توجهی از این گونه سحابیهاست.
جدا از سه گروه سحابیها برخی از سحابیها از ستاره ها تشکیل می شوند. ستاره هایی مانند خورشید در پایان زندگی خود یعنی در مرحله غول سرخی لایه های بیرونی جو خود را به صورت سحابی در فضا می پراکنند. این سحابیها را سیاره نما می نامند. زندگی ستاره های پر جرمتر از خورشید با انفجاری ابرنواختری پایان می یابد و سحابی بزرگ و گسیخته ای از انفجار به جا می ماند که آن را سحابی باقیمانده انفجار ابرنواختری می نامند.
ستاره شناسان چگونه فاصله ستارگان تا زمین را محاسبه می کنند؟
همانطور که می دانید فاصله برخی از ستاره ها تا زمین به هزاران سال نوری می رسد. اگر اخترشناسان می خواستند با محاسبه زمان ارسال و برگشت پرتوهای نوری یا امواج فاصله زمین تا ستاره ها را اندازه بگیرند می بایست هزاران و حتی میلیونها سال منتظر می ماندند.
دانشمندان ریاضیدان راه حل ساده ای به نام اختلاف منظر یافته اند که با این شیوه می توان به راحتی فاصله اجسام دور را محاسبه کرد.
برای فهم بهتر ابتدا مثالی می زنیم : مدادی را مقابل چشمان خود بگیرید. ابتدا چشم چپ را ببندید و با چشم راست به آن نگاه کنید. بعد چشم راست را ببندید و با چشم چپ به آن نگاه کنید. حتما" به نظرتان آمده که مداد چند سانتی متر جابه جا شده است. با همین روش ساده بود که اخترشناسان توانستند شعاع کره زمین و به دنبال آن فاصله ماه و خورشید از زمین را پیدا کنند.
با دانستن فاصله زمین تا خورشید می توان به راحتی فاصله زمین تا ستاره ها را محاسبه کرد.
اخترشناسان از یک ستاره مشخص دو عکس به فواصل 6 ماه از هم می گیرند. وقتی این دو عکس را با هم مقایسه می کنند به نظر می رسد که ستاره چند درجه در آسمان جابه جا شده است. با داشتن فاصله زمین تا خورشید و زاویه ( نصف زاویه ای که به نظر ستاره جابه جا شده ) و به کمک فرمول مثلثاتی ساده می توان فورا" فاصله چند سال نوری از زمین تا این ستاره را محاسبه کرد.
چرا ستاره قطبی را همیشه در یک نقطه ثابت می بینیم؟
زمین دور محوری می چرخد که اگر این محور را به سمت شمال امتداد دهیم در فاصله بسیار دوری به ستاره قطبی خواهیم رسید. فاصله سباره قطبی از منظومه شمسی ما آنقدر زیاد است که مدار گردش زمین دور خورشید مانند چرخش یک نقطه بسیار ریز می ماند. اگر فردی در قطب شمال زمین ایستاده باشد ستاره قطبی را درست بالای سر خود خواهد دید. البته امتداد محور چرخش زمین در فضا ثابت نیست و در یک دوره 26 هزار ساله دور یک دایره می چرخد. بطوریکه تا 2000 سال دیگر امتداد محور چرخش زمین به ستاره دیگری به نام نسر واقع (وگا) ختم می شود و پس از یک دوره 26000 ساله دوباره امتداد محور چرخش زمین با ستاره قطبی فعلی منطبق می شود.
"ویتامین "B5 که به "پانتوتنیک اسید" نیز معروف می باشد، یک ویتامین آنتی اکسیدان محلول در آب است. این ویتامین به سه شکل در می آید و به نام های "پانتنول" و "کلسیم پانتوتنات" نیز نامیده می شود.
همه سلولهای بدن نیاز به ویتامین B5 یا اسید پانتوتنیك دارند. بدن انسان از پانتوتنیک اسید برای تبدیل کربوهیدرات ها ، پروتئین ها و چربی ها به انرژی استفاده می کند.
ویتامین B5 در بدن تبدیل به تركیبی به نام " كوآنزیم A " میشود كه وجود این ماده در بدن برای تبدیل غذا به انرژی لازم است و فقدان آن سبب نارسایی رشد و خاكستری شدن موها و كم خونی می شود.
ویتامین B5 به عنوان ویتامین "ضد استرس" نیز شناخته شده است، زیرا عملكرد غده آدرنال(فوق کلیه) را تقویت كرده و بهبود میبخشد(غده آدرنال عضوی است كه بدن را در مقابل انواع استرسها حمایت میكند).
ویتامین B5 برای عمل عضلات و اعصاب ضروری است و برای حفظ سلامتی سیستم ایمنی بدن، حیاتی است. این ویتامین همچنین در كاهش علایم دردناك روماتیسم مفصلی موثر است. در ضمن این ویتامین رشد و نمو را تسهیل نموده و به سلامت پوست و مو كمك می نماید.
با اینکه کمبود این ویتامین، به دلیل فراوانی آن در غذاها و ساخت آن توسط باکتری های روده، بعید به نظر می رسد، اما خستگی یکی از علائم زودرس و متداول کمبود این ویتامین می باشد.
عملكرد و فواید پنتوتنیك اسید:
* برای رشد، تولید مثل و خیلی از فرایندهای معمول بدن ضروری است.
* شركت در خیلی از واكنش های بیولوژیكی مانند: تولید انرژی، تجزیه اسیدهای چرب و اسیدهای آمینه، ساخت اسیدهای چرب و چربی ها، كلسترول، هورمون های استروئیدی، تولید كو آنزیم A و آنتی اكسیدان سلولی گلوتاتیون
* جزء حیاتی برای ساخت گلبول های قرمز خون
* تولید هورمون های مربوط به استرس و هورمون های جنسی در غدد فوق كلیه(غدد كوچكی كه در بالای كلیه ها قرار دارند)
* مكمل این ویتامین معمولا برای درمان علائم بیماری آلرژی و طیف وسیعی از بیماری های پوستی استفاده می شود.
* تبدیل کربوهیدرات ها، پروتئین ها و چربی ها به انرژی
* برای تولید آنتی بادی های لازم جهت واکسیناسیون علیه کزاز، حصبه و آنفلوآنزای آسیایی لازم است.
* خوردن مکمل این ویتامین(مکمل پنتتئین)، موجب کاهش چربی خون می شود. در فردی با کلسترول خون بالا، خوردن این مکمل، باعث کاهش کلسترول بد(LDL ) و افزایش کلسترول خوب(HDL) می شود.
مقادیر توصیه شده برای دریافت پانتوتنیک اسید
یک میزان RDA رسمی برای ویتامین B5 وجود ندارد، با این حال فقدان پانتوتنیک اسید معمولا با فقدان دیگر ویتامین های گروه B همراه است. لذا به منظور جلوگیری از عدم تعادل ویتامین های گروه B ، B کمپلکس بخورید تا B5 نیز دریافت کرده باشید.
مقدار دریافت روزانه B5 برای بالغین چیزی حدود 7 – 4 میلی گرم در روز می باشد که این مقدار به راحتی در برنامه ی غذایی اکثر افراد عادی یافت می شود.
قرص ها و کپسول های حاوی پانتوتنیک اسید در دوزهای بین 500 – 100 میلی گرمی در دسترس هستند. دوزهای بالاتر در صورت لزوم، ممکن است به وسیله برخی پزشکان، برای درمان های خاص، تجویز شود.
پختن و روش های آماده سازی غذاها و کنسرو کردن موجب تخریب این ویتامین می شود.
کمبود پانتوتنیک اسید
کمبود این ویتامین در بزرگسالان به معنای واقعی وجود ندارد، زیرا در خیلی از منابع غذایی یافت می شود. کمبود این ویتامین ممکن است در افراد الکلی رخ دهد.
با اینکه کمبود این ویتامین، به دلیل فراوانی آن در غذاها و ساخت آن توسط باکتری های روده، بعید به نظر می رسد، اما خستگی یکی از علائم زودرس و متداول کمبود این ویتامین می باشد.
پانتوتنیک اسید در بدن انسان مسمومیت ایجاد نمی کند. اثرات جانبی قابل ملاحظه ای نیز، حتی در دریافت های بالاتر از 10000 میلی گرم (10 گرم) در روز گزارش نشده است.
فقط مقادیر بسیار زیاد، در حد چندین گرم در روز می تواند موجب اسهال شود. غیر از اسهال، واکنش مضری برای مصرف دوزهای بالا گزارش نشده است.
این ویتامین، فواید ویتامین های C ، B6، و B12را در بدن بیشتر و کمبودشان را اصلاح می کند.
در اين مرحله از مولكولهاي NADH و FADH2 توليد شده در مراحل قبلي تنفس سلولي، براي توليد ATP استفاده ميشود.
زنجيرهي انتقال الكترون ميتوكندري، در غشاي داخلي آن قرار دارد.
در پروكاريوتها، محل زنجيرهي انتقال الكترون، غشاي سلولي است.
اين زنجيره، از تعدادي پروتئین غشايي تشكيل شده كه به صورت پمپ عمل ميكنند. الكترونهاي NADH و FADH2 ، با عبور از اين پمپها، باعث رانده شدن يونهاي +H به فضاي بين دو غشا ميشود (انتقال فعال).
در نتيجه به تدريج غلظت +H در اين فضا زياد ميشود و شيب غلظتي در جهت ورود يونهاي به فضاي بين داخلي ميتوكندري (ماتريكس) ايجاد ميشود.
اين يونها از طريق يك پروتئين كانالي به درون ماتريكس وارد ميشوند (انتشار تسهیل شده) و اين پروتئين، در حين عبور ، به ADP، فسفات اضافه ميكند و ATP ميسازد.
الكترونهاي مولكولهاي NADH و FADH2 در نهايت همراه با يونهاي هيدروژن، به مولكولهاي اكسيژن ميپيوندند و مولكولهاي آب توليد ميكنند. اكسيژن پذيرندهي نهايي الكترون است.
به ازاي هر NADH، سه مولكول ATP و به ازاي هر FADH2 ، دو مولكول ATP در زنجيرهي انتقال الكترون ميتوكندري توليد ميشود.
در مجموع از هر مولكول گلوكز، 2 مولكول ATP و 2 مولكول NADH (معادل 6 مولكول ATP) درون سيتوپلاسم تشكيل مي شود. يعني در كل معادل 8 مولكول ATP مولكول پر انرژي درون سيتوپلاسم توليد ميشود.
از هر مولكول گلوكز، درون ميتوكندري، 2 مولكول NADH در مرحلهي تبديل دو پيرووات به استيلكوآنزيم A (معادل 6 مولكول ATP) و 6 مولكول NADH در چرخهي كربس (معادل 18 مولكول ATP ) توليدميشود.
يعني در كل معادل 24 مولكول ATP، مولكول NADH درون ميتوكندري توليد ميشود.
از هر مولكول گلوكز، 2 مولكول (معادل 4 مولكول ATP) تشكيل ميشود كه همه درون ميتوكندري و در چرخهي كربس توليد ميشوند.
از هر مولكول گلوكز، 4 مولكول ATP در سطح پيش ماده توليد ميشود؛ دو تا در گليكوليز و دو تا در چرخهي كربس.
از هر مولكول گلوكز، در مجموع 10 مولكول NADH (معادل 30 مولكول ATP) و 2 مولكول FADH2 (معادل 4 مولكول ATP) و 4 مولكول ATP در سطح پيش ماده توليد ميشود.
يعني در مجموع حاصل يك تنفس معمولي كامل و سوختن كامل گلوكز، 38 مولكول ATP است كه 34 تا در زنجيرهي انتقال الكترون توليد ميشود.
در صورت نبود اكسيژن زنجيرهي انتقال الكترون كارآمد نيست. دراين موقع الكترونهاي NADH به جاي اکسیژن به تركيبات آلي (مانند پيرووات) منتقل ميشوند و +NAD بازسازي ميشود تا گليكوليز تداوم داشته باشد و مقدار اندكي ATP توليد شود.
بازسازي +NAD با استفاده از يك پذيرندهي آلي هيدروژن، تخمير نام دارد. تخمير تجزيهي گلوكز در عدم حضور اكسيژن است.
1- تخمير لاكتيك اسيد
اين تخمير در بعضي باكتريها و قارچها انجام ميشود و براي توليد ماست و انواعي از پنيرها از آن استفاده ميشود.
سلولهاي ماهيچهاي انسان هم در صورت نبود اكسيژن، اين نوع تخمير را انجام ميدهند.
در اين روش، الكترونهاي NADH به پيرووات سه كربني منتقل ميشوند و لاكتات (كه آن هم سه كربني است) را توليد ميكنند. به اين ترتيب +NAD بازسازي ميشود.
2- تخمير الكلي
اين تخمير در مخمرها ديده ميشود كه در نانوايي كاربرد دارند و باعث ورآمدن خمير ميشوند.
در اين روش، ابتدا پيرووات به يك تركيب دو كربني تبديل ميشود (و يك مولكول آزاد ميشود) و الكترونهاي NADH، به اين ترتيب دو كربني اضافه شده، آن را به اتانول تبديل ميكنند. پس +NAD بازسازي ميشود و گليكوليز تداوم مييابد.
گلوکز بوسيله هر دو گروه موجودات هوازي و بي هوازي مصرف مي شود. مراحل ابتدايي تخريب گلوکز که گليکوليز خوانده مي شود براي هر دو گروه يکسان است. در موجودات بي هوازي ، مولکول گلوکز پس از يکسري واکنشهاي تخريبي به فراورده نهايي مبدل مي گردد و پس اين فرآورده در همان شرايط بي هوازي طي مراحل ديگر تخريب مي شود و يا به مصرف مي رسد. در موجودات هوازي ، مرحله گليکوليز بطور مشابه با شرايط بي هوازي طي مي شود و پس فرآورده بدست آمده که پيرووات است در شرايط هوازي تخريب نهايي حاصل کرده، CO۲ و آب و انرژي توليد مي کند.
● گليکوليز
گليکوليز در سيتوپلاسم ياخته رخ مي دهد و شامل ۱۰ واکنش شيميايي پي در پي است. گليکوليز بيان ساده اي از استخراج انرژي شيميايي نهفته در ترکيبات آلي مواد غذايي است که طي آن در شرايط بي هوازي يک مولکول گلوکز به دو مولکول پيرووات مبدل مي شود. گليکوليز از کلمه يوناني گليکو به معني شيرين و ليز به معني کاستن و از دست دادن گرفته است. واکنشهاي گليکوليز به صورت خطي پيش مي روند. راه گليکوليز به نام دو دانشمندي که در مشخص کردن اين راه بسيار کوشيدند به راه امبدن ميرهوف موسوم است.
▪ مراحل گليکوليز
در نخستين بخش ، گلوکز با مصرف دو مولکول ATP فسفريل دار و فعال مي شود و به فروکتوز ۱ ۶ دي فسفات مبدل مي گردد. در مرحله دوم مولکول اخير به دو مولکول سه کربني به نامهاي گليسرآلدهيد ۳ فسفات و دي هيدروکسي استون فسفات تجزيه مي شود و اين دو مولکول ايزومر يکديگرند. در مرحله سوم ، گليسرآلدهيد و ۳ فسفات اکسيده شده و با از دست دادن هيدروژن خود به اسيد ۱ ۳ دي فسفوگليسريک مبدل مي شود.
هيدروژن آزاد شده براي احياي NAD و ايجاد ۲ مولکول NADH بکار مي رود. در مرحله چهارم که مرحله توليد انرژي است اسيد دي فسفاته حاصل با از دست دادن فسفات خود و فسفريل دار کردن ADP موجب ساخته شدن ۴ مولکول ATP شده و خود سرانجام به اسيد پيرويک تبديل مي شود.
● تخمير الکلي
سرنوشت پيرووات در شرايط بي هوازي کاملا متفاوت با شرايط هوازي است. در صورتي که شرايط بي هوازي باشد، پيرووات ابتدا تحت اثر آنزيم دکربوکسيلاز گروه کربوکسيل خود را به صورت CO۲ از دست مي دهد و به استالدئيد تبديل مي شود در مرحله بعد ، استالدئيد تحت تاثير آنزيم الکل دهيدروژناز به اتانول مبدل مي شود. فرآيند تبديل پيرووات به اتانول و CO۲ تخمير الکلي ناميده مي شود. در تخمير الکلي تعداد ATP توليد شده مانند راه گليکوليز است.
● راه پنتور فسفات
يکي ديگر از راههاي تخريب گلوکز راه پنتوز فسفات است که به علت ايجاد قندهاي پنج کربني و ساير قندها اهميت دارد. اين راه فرآيندي است که طي آن از يک سو گلوکز ماهيت خود را از دست مي دهد و تخريب مي شود و از سوي ديگر فراورده نهايي آن توليد NADPH و سنتز ساير قندها بويژه ريبوز است. راه پنتوز فسفات هم به عنوان فرايندي از تخريب کربوهيدرات و هم به عنوان فرايندي در سنتز برخي از قندها به شمار مي آيد اين راه نيز مانند گليکوليز در سيتوپلاسم ياخته رخ مي دهد.
● تخريب ساير قندها
قندهاي ديگر مانند گالاکتوز ، فروکتوز ، مانوز طي يک سري واکنشهاي اختصاصي به متابوليسمهاي راه گليکوليز تبديل شده و سپس تشکيل پيرووات را مي دهند.
● چرخه کربني
اسيد پيرويک حاصل از گليکوليز در حضور اکسيژن کافي ابتدا کربوکسيل زدايي و سپس اکسيد مي شود و با اتصال به يک مولکول آلي پيچيده به نام کوآنزيم A ، ترکيب دو کربني جديدي به نام استيل کوآنزيم A را مي سازد. اين واکنش پيچيده است و به وجود حداقل ۵ کوآنزيم يا کوفاکتور نياز است. استيل کوآنزيم A بوجود آمده با داشتن آرايش فضايي مناسب موجب شروع واکنشهاي چرخه کربني مي شود. بطور کلي چرخه کربني شامل چهار مرحله اکسايش است. چرخه کربني مستلزم ده واکنش آنزيمي است که طي آن در جمع بندي کل سه ترکيب اصلي CO۲، NADH و FADH۲ و مقدار کمي انرژي به صورت GTP بدست مي آيد.
● سيستم انتقال الکترون وفسفريلاسيون اکسيداتيو
در موجودات هوازي در آخرين مرحله ، آنزيمها و فرآورده ها يا سوبستراهاي کاهنده حاصل از چرخه کربني با يک سري از مواد انتقال دهنده الکترون همراه مي شوند که در نتيجه اين همراهي و همبستگي ، الکترونهاي حاصل از اکسايش اين سوبستراها موجب احياي مواد ناقل الکترون که زنجيره وار برحسب پتانسيل رودکس قرار گرفته اند مي گردند و در پايان زنجيره اکسيژن را که گيرنده نهايي الکترون است احيا کرده و آب را تشکيل مي دهند.
احيا و سپس اکسيده شدن دوباره هر يک از ناقلين سيستم انتقال الکترون موجب رها شدن انرژي مي گردد که اين انرژي صرف سنتز ATP مي شود. اين فرآيند سنتز ATP را که با استفاده از انرژي رها شده از جريان الکترون از طريق سيستم انتقال الکترون (که انتهاي آن اکسيژن قرار دارد)، فسفريلاسيون اکسيداتيو با فسفريل دار شدن اکسايش مي گويند.
از صفات غالب وابسته به X در انسان را می توان بیماری ریکتز را نام برد که موجب کاهش میزان فسفات خون می شود .الل غالب موجب بروز بیماری می گردد. از ازدواج مرد مبتلا با زن سالم تمام دختران آ نها مبتلا و تمام پسران سالم خو اهند بود . از آنجا که در حالات طبیعی کروموزوم X پدری به دختر انتقال می یابد بنابراین بیماری ریکتز از پدر به تمام دختران منتقل شده است ولی از ازدواج زن هترو زیگوت با مرد سالم نیمی از پسران و دختران مبتلا ونیم دیگر سالم خواهند بود .
3) در باکتری ها، نقش کلروپلاست را چه چیز ایفاء می کند؟ غشای سلولی تخصص یافته
4) هریک از مراحل فتوسنتز در کدام قسمت کلروپلاست انجام می شود؟
مرحله 1 (نوری) در تیلاکوئیدها، مرحله 2 (نوری) در تیلاکوئیدها ولی محصول به استروما رها می شود، مرحله 3 (تاریکی) در استروما
5) چرا گیاهان سبز دیده می شوند؟
6) چون رنگ سبز نور مرئی را، جذب نکرده و به چشم باز تابش می کنند.
7) استفاده از کلروفیل ها و کاروتنوئیدها بطور همزمان برای یک گیاه چه فایده ای دارد؟
کاروتنوئیدها سبز را هم جذب می کنند و کلاً میزان جذب نور بالا می رود.
8) کدام طول موج تقریباً توسط رنگیزه ها جذب نمی شود؟زرد
9) کلروفیل ها کدام نورها را بیشتر جذب می کنند؟ قرمز، آبی، بنفش
10) کاروتنوئیدها کدام نورها را بیشتر جذب می کنند؟ آبی و سبز
11) تفاوت های فتوسیستم I و II را بنویسید.
الف- طول موجی که حداکثر جذب را دارند، یا 680 نانومتر است یا 700 نانومتر. ب- نوع کلروفیل a ویژه آنها متفاوت است.
12) چرا در کلروفیل a ویژه، الکترون، برانگیخته می شود؟
چون الکترون مدار آخر منیزیم وسط کلروفیل a ویژه، بر اثر تابش نور و انرژی آن برانگیختهمی شود و فتوسیستم ها را ترک می کند.
13) خلاء الکترونی فتوسیستم II و I به ترتیب چگونه جبران می شود؟
کمبود الکترونی PII توسط آب و PI توسط PII جبران می شود.
14) دهنده اولیه و گیرنده نهایی الکترون در زنجیره انتقال الکترون در کلروپلاست چه می باشند؟دهنده= آبگیرنده= NADP+
15) پمپ پروتون زنجیره انتقال الکترون در کلروپلاست کدامست و انرژی خود را چگونه تأمین می کند؟ پروتئین وسطی، از نشت انرژی الکترون های در حال حرکت در زنجیر انتقال الکترون.
16) دلایل افزایش پروتون در تیلاکوئید چیست؟ الف- شکسته شدن H2Oب- عملکرد پمپ پروتون
17) پروتئین کانالی که خاصیت آنزیمی هم دارد، انرژی لازم برای ساخت ATP را از کجا تأمین می کند؟ از شیب غلظت پروتون ها و نیروی حاصل از ازدحام آنها
18) محصول زنجیره اول و زنجیره دوم انتقال الکترون در کلروپلاست به ترتیب چه می باشد؟
زنجیره اول ATP، زنجیر دوم NADPH
19) موجودات فتوسنتز کننده به چند روش، دی اکسید کربن را تثبیت می کنند؟
تبادل
لینک هوشمند
برای تبادل
لینک ابتدا ما
را با عنوان
زیست -زمین و آدرس
biogeology.LXB.ir لینک
نمایید سپس
مشخصات لینک
خود را در زیر
نوشته . در صورت
وجود لینک ما در
سایت شما
لینکتان به طور
خودکار در سایت
ما قرار میگیرد.